Tong Li1,
Shan Jiang1,
Zi-Jun Wang1,
Li-An Huang2 ![]()
For correspondence:- Li-An Huang Email: huanglian09@gmail.com Tel:+8657786699816
Received: 17 August 2015 Accepted: 3 June 2016 Published: 31 July 2016
Citation: Li T, Jiang S, Wang Z, Huang L. Withaferin A promotes proliferation and migration of brain endothelial cells. Trop J Pharm Res 2016; 15(7):1487-1492 doi: 10.4314/tjpr.v15i7.19
© 2016 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To investigate the effect of withaferin A (WFA) on the proliferation and migration of brain endothelial cells.
Methods: BALB-5023 mouse microvascular cells were treated with a range of withaferin A (WFA) concentrations from 10 to 100 ng/mL. Dojindo’s CCK-8 cell proliferation kit was used for the analysis of cell proliferation. Transwell cell culture inserts were used to determine the migration potential of WFA-treated endothelial cells. Absorbance was measured at 450 nm on an enzyme-linked immunosorbent (ELISA) reader.
Results: The results revealed a significant increase in the proliferation and migration of endothelial cells following treatment with a low concentration (30 ng/mL) of WFA compared with the higher concentration (> 10 ng/mL). The effect was further enhanced when WFA was used in combination with soluble Fas ligand (sFasL). Autocrine signaling of vascular endothelial growth factor (VEGF) by endothelial cells was significantly increased following treatment with WFA or in combination with sFasL. WFA increased the ex
Conclusion: Thus, WFA promotes the proliferation and migration of endothelial cells through increase in the ex
Introduction
At present the leading cause of morbidity and mortality throughout the globe is cardiovascular diseases and stroke. Endothelial cells form the covering of the vascular lumen of peripheral organs and the central nervous system. These cells produce and release substances that relax or constrict the blood vessels, which may contribute to the development of vascular failure [1]. To maintain the necessary supply of metabolic and functional requirements to various tissues formation of new blood vessels or angiogenesis has a vital role. Angiogenesis involves the proliferation and migration of endothelial cells [2]. The immune compromised mice with stroke on administration of human cord blood-derived CD34+ cells develop neovascularization in the ischemic zone and provide a favourable environment for neuronal regeneration [3]. It is reported that circulating CD34+ endothelial progenitor cells participate in neovascularization in ischemic tissues [4,5]. But the outgrowth of pre-existing vasculature is assumed to be indispensable in the postnatal development of neovessels. Therefore the proliferation and migration ability of endothelial cells is vital for the recovery of vascular diseases, including cardiovascular diseases and stroke.
Withaferin A [1], a steroidal lactone is isolated from Withania somnifera, the extracts of which are used in traditional East Indian medicine [6]. Among the withanolides isolated from W. somnifera, withaferin A is present as a dominant compound [7,8]. Withaferin A has anti-angiogenic effect [6, 9] at micromolar doses and inhibits soft tissue sarcoma growth and local recurrence in xenograft experiments [10]. It is reported to have proapoptotic and anti-tumor activity in breast and prostate cancers [11-13]. Withaferin A acts on NF-Kb, BCL-2, FOXO3A, Hsp90, phosphorylated STAT3 and annexin II [12,14-18]. Taking cue from the literature, we designed an experiment to study the effect of withaferin A on suppression of brain tumor growth in a nude mice model with an attempt to develop potent therapeutic agent.
Methods
Cell line and culture
The mouse brain microvascular endothelial cell line, BALB-5023 was purchased from the American Type Culture Collection (Manassas, VA, USA). The cells were grown in complete medium consisting of DMEM Gluta MAX, supplemented with 1 % penicillin/streptomycin and 10 % FCS (Gibco-BRL, Melbourne, Australia).
Endothelial cell proliferation assay
For effects on cell proliferation, Dojindo’s CCK-8 cell proliferation kit (Dojindo Molecular Technologies, Japan) was used. Briefly, 1 × 105 cells in 100 μL complete medium were seeded into each well of a 96-well plate. After 24 h, different concentrations of WFA (Chinese National Institute for the Control of Pharmaceutical and Biological Products, Beijing, China) or soluble FasL (Sigma, St. Louis, MO, USA) were added to each well containing endothelial cells for 48 h. 20 μL of CCK-8 solution (5 mg/mL; Dojindo, Kumamoto, Japan) was added into each well after incubation for 48 h. The cells were then incubated for 4 h more. A Multiskan Go spectrophotometer (Thermofisher Scientific, USA) was employed to measure the cell proliferation at 450 nm.The proliferation index was calculated using the equation 1.
(OD450 in the presence of WFA - OD450 in the blank control)/OD450 in the blank control × 100 %………………………………………………1
Endothelial cell migration assay
Transwell cell culture inserts were used to determine the migration potential of WFA treated endothelial cells. The upper chambers of the inserts were seeded with 2 × 105 cells in 200 mL serum-free DMEM whereas the lower chamber was filled with 750 mL of DMEM containing 20 % FBS as a chemoattractant except in control wells which contained serum-free DMEM in both upper and lower chambers. After 24 h of incubation in a humidified incubator with 5 % CO2 the non-migrated cells in the upper chamber were swabbed off. The plates were fixed, stained, and then observed under an inverted fluorescent microscope. Five field views were randomly counted in triplicates and averaged.
ELISA for VEGF
In 6-well plates, endothelial cells were treated for 72 h with either WFA or WFA and soluble FasL. The supernatant was collected and stored at -80 oC. For VEGF, ELISA (R&D Systems, Minneapolis, MN, USA) was performed as per the manufacturer's protocol. To each well 100 μl assay diluents, followed by 100 μl standard, control or cell culture supernatant was added. The plates were incubated at room temperature for 2 h followed by aspiration and washing. Thereafter, 100 μL conjugated secondary antibody was added into each well for 2 h at room temperature. After washing, 100 μL of substrate solution was added to each well for 30 min at room temperature. Finally, 100 μL of stop solution was added and the absorbance was read at 450 nm on an ELISA reader (Bio-Rad, Hercules, CA, USA) within 30 min. All readings were performed in triplicates.
Western blotting for Fas protein
The cells treated with WFA alone or combined with soluble FasL for 72 h were washed twice in PBS followed by addition of Lysis buffer (50 mM Tris-HCl pH 7.4, 137 mM NaCl, 10 % glycerol, 100 mM sodium vanadate, 1 mM PMSF, 10 mg/ml aprotinin, 10 mg/mL leupeptin, 1 % NP-40, and 5 mM cocktail). Bicinchoninic acid assay (BCA) method was used to determine protein concentration. Equal amounts of protein were loaded and resolved by electrophoresis on a 10 % polyacrylamide gel. The semi-dry method was used to transfer proteins onto a polyvinylidene difluoride (PVDF) membrane which was then blocked with 5 % non-fat dry milk overnight. After Tris-buffered saline Tween-20 (TBST) washing, membrane was incubated for 2 h with anti-Fas (dilution 1:2,000; KeyGen Biotech, Nanjing, Jiangsu, China) and then washed again with TBST before incubation with secondary antibodies for 2 h. Antigen was detected using enhanced chemiluminescence (Pierce Biotechnology, Inc, Rockford, IL, USA). All the samples were normalized to β-actin.
Statistical analysis
Each assay was performed in triplicate. The data are expressed as mean ± standard deviation (SD). SPSS 16 software was used for all statistical analyses. One-way analysis of variance (ANOVA) was used to determine significant differences. Statistical significance was set at p < 0.05.
Results
Proliferation of endothelial cells stimulated by low-dosage WFA
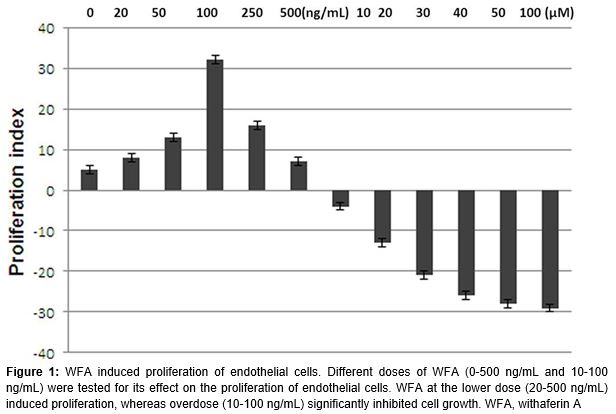
The endothelial cells were treated with a range of WFA concentrations and analysed by CCK-8 cell proliferation assay kit. The results revealed that WFA stimulated the proliferation of the endothelial cells significantly at a low-dosage, ranging from 0.025 to 0.25 ng/mL, compared to the control cells (p < 0.05). However, treatment of the cells with WFA at higher dosage (> 100 ng/mL), resulted in the cell impairment (). Thus, 0.25 ng/mL WFA with sFasL was used to analyse the combined effects.
Effect of sFasL on the WFA stimulated proliferation of endothelial cells
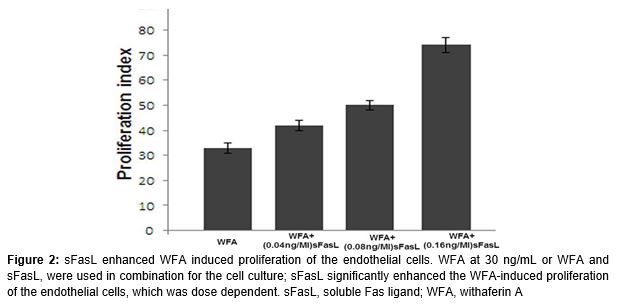
Treatment of the endothelial cells with WFA at 0.25 ng/mL induced the cell division. sFasL at the concentration of 0.16 ng/mL also induced the proliferation of endothelial cells through Fas-FasL pathway. However, the proliferation of endothelial cells was significantly higher on treatment with WFA and sFasL combination compared to WFA alone (p < 0.05; ).
WFA combined with sFasL enhances the migration of endothelial cells
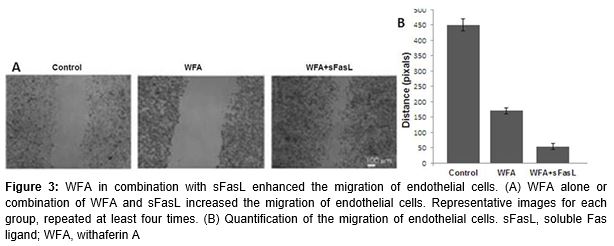
The transwell cell culture inserts were used to study the effect of WFA on the endothelial cell migration. The results revealed that WFA promoted the proliferation of endothelial cells. Endothelial cell migration is also essential for angiogenesis. In the present study, WFA could also enhance the migration of endothelial cells, which is more evident if combined with sFasL (p < 0.05; ).
Fas expression increases upon stimulation of WFA
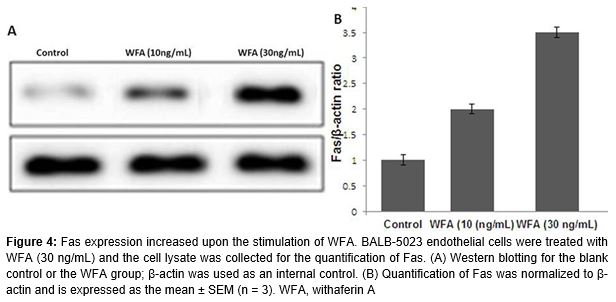
WFA and sFasL could promote the proliferation and migration of endothelial cells. In addition, WFA also increased the expression of Fas on endothelial cells (p < 0.05; ).
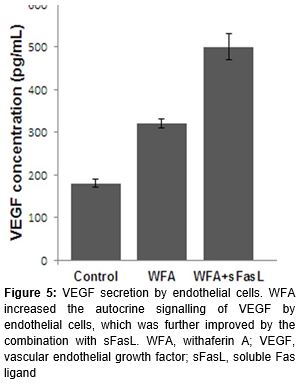
WFA promotes the autocrine signaling of VEGF by endothelial cells
The proliferation and migration of endothelial cells were mediated by VEGF. Accordingly, the endothelial cells secreted significantly more VEGF upon the stimulation of WFA. sFasL combination further increased the autocrine signaling of VEGF ().
Discussion
It is reported that WFA exhibits anti-angiogenic effect [12,15] at micromolar doses and inhibits soft tissue sarcoma growth and local recurrence in xenograft experiments [16]. Withaferin A can also inhibit the NO production by modulation iNOS.
The present study demonstrates that WFA significantly enhanced the proliferation of the endothelial cells at a lower concentration (30 ng/mL). However, at a concentration > 10 µM WFA exhibited reverse effect on the proliferation of endothelial cells in the physiological condition. It was observed that overdose of WFA had a harmful effect on the endothelial cells. In addition to enhanced cell proliferation, WFA also increased the migration of the endothelial cells. Both the enhanced cell proliferation and migration were significantly promoted by sFasL. The effect of WFA on Fas-FasL pathway in the endothelial cells was also examined. It is reported that Fas-FasL ligation induces proliferation of the endothelial cells by expressing the Fas-associated death domain protein and the Flice-like inhibitory protein (FLIP).
FLIP recruits and activates the downstream molecules TNF-receptor-associated factor and nuclear factor κB (NF-κB). This down regulation finally induces the proliferation of the cells. Among the downstream molecules of the Fas-FasL pathway NF-κB plays a vital role. In the present study, WFA directly up-regulated the expression of Fas, suggesting the underlying mechanisms of WFA on endothelial cells.
Conclusion
WFA promotes the proliferation and migration of endothelial cells and the effect is enhanced by combining it with sFasL. The effect of WFA on the endothelial cells is dependent on the increased expression of Fas and enhanced secretion of VEGF.
References
Archives


News Updates